Cellular Mechanisms of the Color Constancy in the Fish Retina
Elena M. Maximova and Vadim V. Maximov
Institute for Problems of Information Transmission,
Russian Academy of Sciences, 101447, Moscow, Russia
 A paper version of the poster was presented
A paper version of the poster was presented
at the meeting in honour of John Lythgoe
"The Ecology of Vision"
Oristano, Italy (6th - 10th April, 1994)
*
Abstract. We supposed that color-coding double-opponent ganglion cells
can display color constancy, for their receptive field (RF) surround might
supply the center with the information about the ambient illumination,
and its opponent influence on the center might well make necessary scaling
for the central signals.
Responses of a certain type of color-coding ganglion cells to flat colored
stimuli moving across the RF center were recorded from their axonal terminals
in tectum opticum. All scene projected onto the RF was illuminated with light
whose spectral composition was changed in a wide range. Under white illumination
blue-green stimuli elicited OFF central response while the red ones elicited ON-OFF
central response. When the RF surround was colored white, these types of responses
remained unchanged while the incident light was varied from red to blue altering
the spectral content of the light reflected from the stimuli. When the RF surround
was covered with black velvet (i.e., in lack of information about the illumination),
one could change the pattern of the central response by varying spectral composition
of the illumination. Moreover, covering the surround with colored papers we
transformed the pattern of the central response in a predictable manner even
under white illumination. This means that the constantly illuminated surround
signals the cell about the character of illumination.
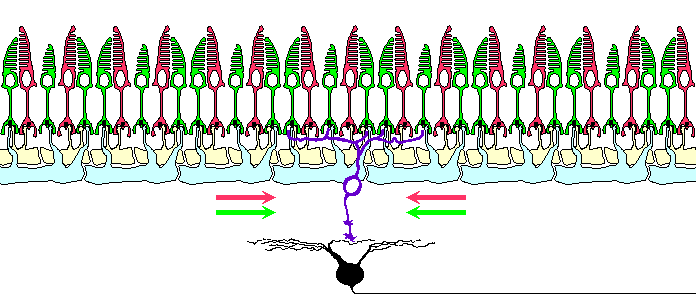
Horizontal cells are known to organize the surround of the ganglion cells. Thus,
owing to their prolonged polarization during illumination, horizontal cells can
realize some color constancy mechanisms just at the very beginning of the visual
pathway.
Preliminary reports of these findings have been made elsewhere
in Russian1
and in English2.
Table of contents
Introduction
In natural environments illumination varies in a wide range, so varies
spectral content of the light reflected from the objects. Nevertheless,
animals do recognize surface colors correctly, irrespective of the illumination.
This ability is the fundamental property of color vision known as
color constancy.
One of the possible mechanisms of the color constancy consists in discounting
the illumination. It requires to determine color of the prevailing illumination
what visual system can do even if there is no illuminant itself in the visual field.
In this case visual system uses some indirect features of the illumination.
For example, it is sufficient to have some white object in the visual field
to estimate the illumination color.
Mechanisms of the color constancy may be of different levels of complexity,
the simplest ones, that do not need memory or learning, may be of retinal origin.
Color-coding opponent ganglion cells of the fish retina were chosen as objects
for the following reasons:
- fishes are known to have good color vision,
- color constancy was found in behavioral experiments
with fishes3,
- opponent code permits to judge what color the ganglion cell perceives,
- surround of the receptive field might supply the center with the information
about the ambient light,
- its opponent influence on the center might well make necessary scaling
for the central signals.
Materials and Methods
Responses of retinal ganglion cells were recorded from their axonal
terminals in the tectum opticum of Cyprinus carpio L. and
Carassius carassius (L.)
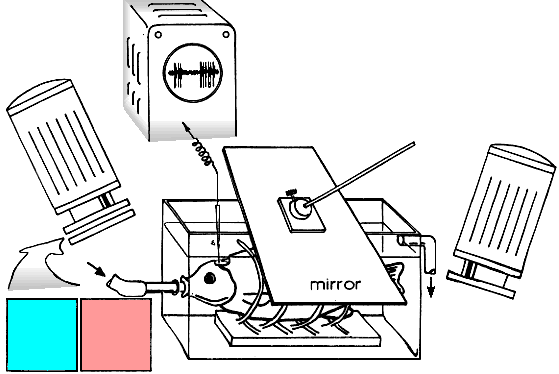

The receptive fields (RF) of the unit was mapped by means of a flashing light-point
and moving black or white spots, stripes or edges. Then, a special device to project
moving stimuli into the RF was positioned at the visual field and the right projection
was adjusted with a mirror.
All the scene was illuminated with two independent sources. Their intensities and
spectral contents were varied in a wide range.
While investigating the color constancy stimuli (sheets of colored papers) were
moved with a constant velocity across the RF center on a velvet-black background,
the RF surround being covered with a stationary white or black screen.
The spike activity of the unit was recorded in response to introduction
(ON response) and withdrawal (OFF response) of the stimulus.
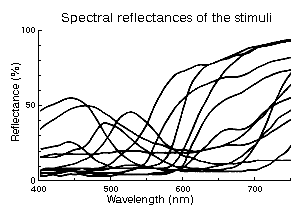

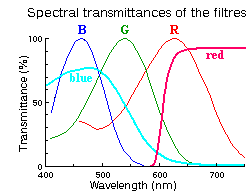
 The relative efficiencies (quantum catches) of the papers under white incandescent
lamp illumination were calculated for each cone type from
The relative efficiencies (quantum catches) of the papers under white incandescent
lamp illumination were calculated for each cone type from
� spectral sensitivities of the cones,
� spectral reflectances of the papers and
� the spectrum of the illuminant.
The efficiency of perfect white diffuser was taken as 100%.
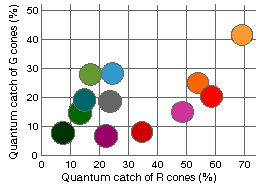
Positions of the papers relative efficiency in the cyprinid color space
are shown in the RG-plane projection to the right.
Stratification of Tectal Activity
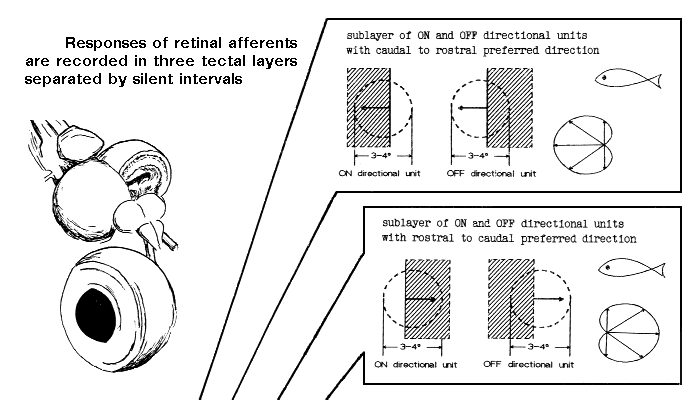
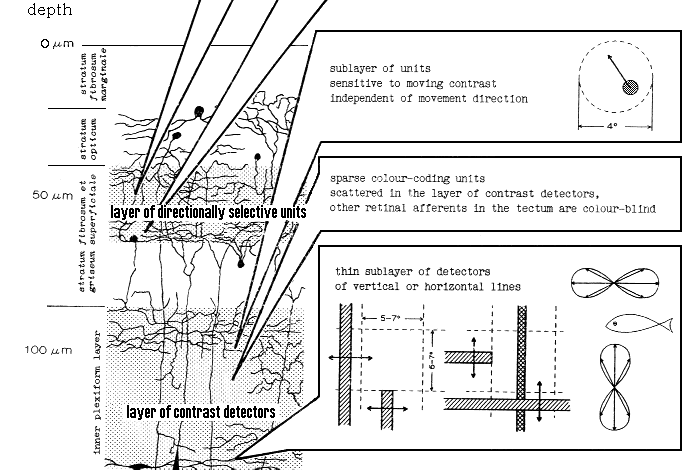
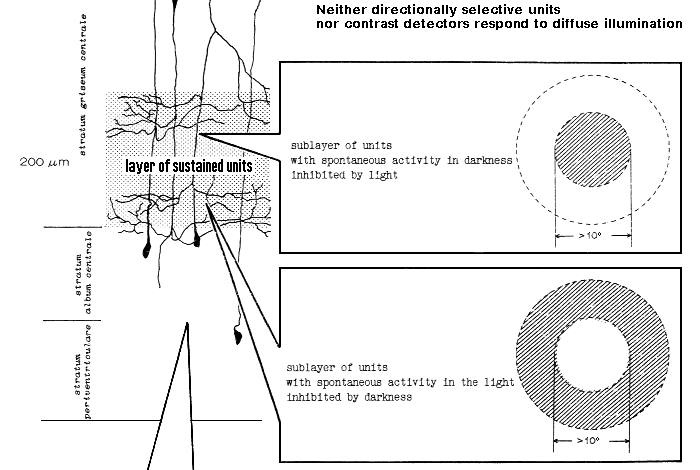

The same types and the same sequences of retinal afferents were observed in the tectum
of the pike4 and some
marine5 fishes:
golden gray mullet |
Liza aurata (Risso) / Mugil auratus (Risso) |
wrasses |
Symphodus sp. / Crenilabrus sp. |
red mullet |
Mullus barbatus ponticus Essipov |
high-body pickarel |
Spicara smaris (Linnaeus) |
Black Sea scad |
Trachurus mediterraneus ponticus Aleev |
Properties of the detectors of vertical and horizontal lines were thoroughly investigated
later and published
elsewhere6,7.
General Properties of Color-Coding Units
Unlike a well-known wide variety of color-coding ganglion cells in the fish retina,
only one type of color-dependent responses is recorded from retinal afferents
projected into the tectum opticum.
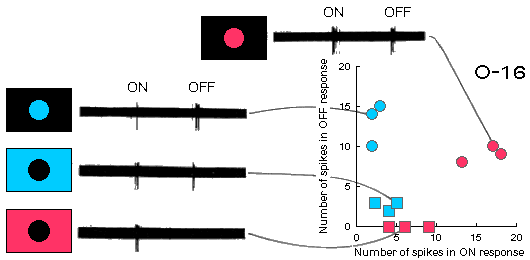 Fig.G1 (above). Spike discharges of the color-coding unit O-16 to long-wave
and short-wave light steps presented into its RF center and surround.
Fig.G1 (above). Spike discharges of the color-coding unit O-16 to long-wave
and short-wave light steps presented into its RF center and surround.
One can see that these responses differ a bit from the classic double color-opponent
ones: there are some extra OFF-response to red illumination of the RF center and
an extra ON-response to blue illumination of the RF surround.
Shown in the panel on the right side of the figure are responses represented
in a graphics form. Three recordings were made for each type of stimulation
in the experiment. Separate points in the space of responses correspond to different
individual records, thus illustrating an experimental scatter.
 Fig.G2. Spike discharges of the color-coding unit O-16 in response to introduction (ON)
and withdrawal (OFF) of sheets of colored papers in its RF center.
Fig.G2. Spike discharges of the color-coding unit O-16 in response to introduction (ON)
and withdrawal (OFF) of sheets of colored papers in its RF center.
Color-coding units receive inputs from red and green cones as revealed
in color-matching experiments.
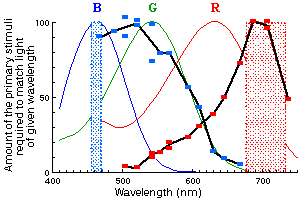
Fig.G3 (to the right). Two color-matching distribution coefficients (shown in black)
obtained for the color-coding unit O-103.
The points indicate relative intensities of long-wave and short-wave lights
(dashed spectral bands) in color mixtures, indistinguishable from certain
monochromatic light (abscissa), the absence of reaction in response to the color
substitution being a criterion of color match.
This type of retinal afferents demonstrates clear color-coding properties,
their ON response being an indicator of the relative amount of a long-wave radiation
in the light, reflected from the stimulus.
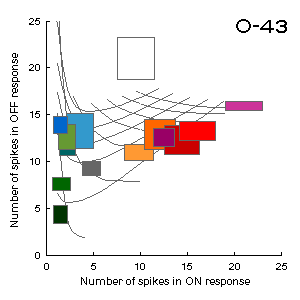
Fig.G4 (to the left). The correspondence between the space of responses (ON-OFF)
of the color-coding unit O-43 and the RG-plane of the cyprinid color space.
Responses of the unit were recorded to presentations of various colored sheets
of paper in black surround under white illumination. Dimensions of the rectangles
at the figure correspond to dispersions of the experimental points.
The curvilinear grid represents the distortion of the color space
(see the RG-plane projection in section Materials and Methods for comparison).
Color Constancy
In lack of information about the color of the illumination (in black surround)
responses of the color-coding units are determined by the light reflected from
the surface presented in the RF center. So, these responses change essentially
when the spectral content of the illumination varies.
When the information about the illumination is available (in white surround)
the unit response keeps rather constant, it does not depend on the color of
illumination and is determined exclusively by the surface color of the stimulus.
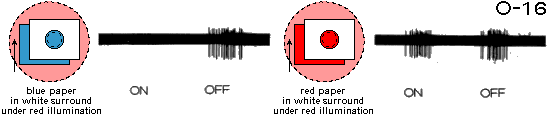 Fig.C1. Spike discharges of the color-coding unit O-16 in response to introduction (ON)
and withdrawal (OFF) of sheets of colored papers in its RF center
(see Fig.G2 in section General Properties of Color-Coding Units for comparison).
Fig.C1. Spike discharges of the color-coding unit O-16 in response to introduction (ON)
and withdrawal (OFF) of sheets of colored papers in its RF center
(see Fig.G2 in section General Properties of Color-Coding Units for comparison).
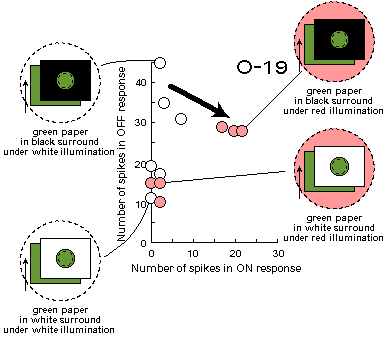
Fig.C2 (to the right). Cell responses to the green stimulus under white
and red illumination in black and white surround.
One can see that responses change (arrow) after switching on an additional red light source,
when the surround is black, and do not change, when it is white.
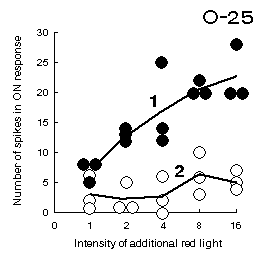
Fig.C3 (to the left). An increase in number of spikes in the ON response
of the unit O-25 to introduction of the green stimulus into its RF center
with reddening the illumination when the RF surround is black (curve 1)
and the relative constancy of the response to the same stimulus
when the RF surround is white (curve 2).
During these experiments, human observer also perceives the stimulus as red at large intensities
of the additional red illumination, if the surround is covered with a black screen and there are
no information about the illumination, but sees the stimulus as green, if the surround is white.
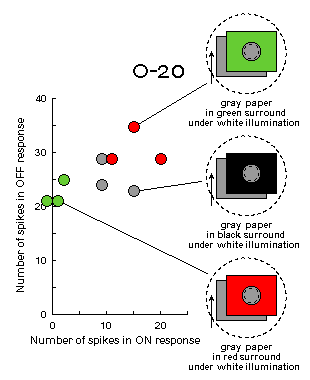
Fig.C4 (to the right). Experiments on simultaneous color contrast.
Under white illumination when the RF surround is covered with a green screen,
ganglion cell considers the illumination to be green and responds to the gray
stimulus as to the red one - its OFF-response increases. This is a well known
phenomenon of simultaneous color contrast.
Opposite illusion is observed when the RF surround is covered with a red screen.
Note, that neither surrounds nor illumination change in the course of stimulation
in these experiments. Only gray paper moves across the RF center on a velvet-black background.
Conclusion
Tectum opticum (with its only type of color-coding retinal afferents) is hardly the
very center where color information is processed. Nevertheless, responses of the
color-coding unit, recorded in the tectum, can reflect the events occurred in the retina.
 Our results suggest that the pattern of response of color-coding opponent ganglion cells
is not determined solely by the spectral content of the light reflected from objects
presented in their RF center, but rather by the surface color of these objects,
provided the information about the illumination is available. Thus, the simplest
mechanisms of the color constancy exist at the retinal level.
Our results suggest that the pattern of response of color-coding opponent ganglion cells
is not determined solely by the spectral content of the light reflected from objects
presented in their RF center, but rather by the surface color of these objects,
provided the information about the illumination is available. Thus, the simplest
mechanisms of the color constancy exist at the retinal level.
Horizontal cells have been shown to organize the surround of the ganglion cell receptive
field8. Artificial sustained polarization
of the horizontal cells, as well as sustained illumination of the ganglion cell RF surround,
modifies the response of the ganglion cells to stimuli presented in their RF
center9. So, it is reasonable to suggest
horizontal cells to be responsible for realization of some color constancy mechanisms just
at the very beginning of the visual pathway.
Literature
1.
E.M.Maximova, A.M.Dimentman, V.V.Maximov, O.Yu.Orlov.
Electrophysiological investigation of the colour constancy in retina.
In: Mechanisms of Action of the Receptor Elements of Sensory Organs -
Proceedings of The 2nd All-Union Symposium on Mechanisms of Reception
(Pushchino, 1971), "Nauka", Leningrad, 1973, pp. 75-80 (in Russian)
2.
E.M.Maximova. Cellular mechanisms of colour constancy. (Presented at
The 2nd International Congress of C.I.A.N.S. Prague, 1975),
Activ. nerv. sup. (Praha), 19, 3:199-201, 1977
3.
A.M.Dimentman, A.Ya.Karas, V.V.Maximov, O.Yu.Orlov. Constancy of object colour
perception in Cyprinus carpio. Pavlov J. Higher Nerv. Act. 22,
4:772-779, 1972 (in Russian)
4.
G.M.Zenkin, I.N.Pigarev. Detector properties of the ganglion
cells in the pike retina. Biofizika 14:763-772, 1969 (in Russian)
5.
E.M.Maximova, O.Yu.Orlov, A.M.Dimentman. Investigation of the visual
system of some marine fishes. Voprosy Ichtiologii 11, 5:893-899,
1971 (in Russian)
6.
E.M.Maximova, V.V.Maximov. Detectors of oriented lines in the visual system
of the crucian carp Carassius carassius. J. evol. Biochem. & Physiol.
17, 5:519-525, 1981 (in Russian)
7.
E.M.Maximova. Colour and spatial properties of detectors of oriented
lines in the fish retina. Iugoslav. Physiol. Pharmacol. Acta 34,
2:351-357, 1998
The paper is available in PDF format at
http://147.91.239.50/ippa/issues/34(2)/IPPA1998342_351_356.pdf (241K)
8.
E.M.Maximova. Effect of intracellular polarization of horizontal cells
on ganglion cell activity in the fish retina. Biofizika 14, 3:537-544,
1969 (in Russian)
Translated in: Neuroscience Translations No.11, pp.114-120, 1969-1970
9.
E.M.Maximova, V.V.Maximov. The role of horizontal cells in
organization of ganglion cells concentric receptive fields in the fish
retina. In: Visual Information Processing and Control of Motor Activity
- Proceedings of the International Symposium (Sofia,1969), Bulgarian
Acad. of Sci., Sofia, 1971, pp. 31-39 (in Russian)
Questions and comments to
Back to IITP Laboratory 8 Activity Page
Back to IITP Electronic Posters Page
Back to IITP Projects Page ( IITP russian Projects Page )
Last Update: 14 February, 2000