This page contains original electronic versions of the figures to the paper by E.M. Maximova et al.,
as well as some supplementary images of ganglion cells traced with DiI in the fish retinas that were not included in the paper.
The paper will be published in Russian in Sensory Systems, 2006, 20 №4.
The page may also be considered as an electronic version of the presentation
"Morphology of putative direction-selective ganglion cells traced with DiI in the fish retina as compared with those of mammals"
by E. Maximova at the International Biophysics Workshop "Academician Radoslav K. Andjus" on Imaging in Neurosciences and Beyond, Sveti Stefan (Montenegro), September 23-30, 2006.
Abstract Visual processing starts already in the retina.
In the fish there are many types of ganglion cells (GCs) specified in detection of
different properties of visual objects such as orientation, size, direction
of movement, sign of contrast and so on (Maximova et al., 1971; Maximov et al., 2005).
These specific abilities are determined by definite wiring in the inner plexiform layer (IPL)
where GCs receive input signals from different bipolar and variety of amacrine
cells. Therefore GCs of different physiological functions (detectors of different types)
would differ morphologically, in particular, their dendrites could be expected to differ in shape,
size, mode of stratification (Rockhill et al., 2002; Mangrum et al., 2002). At the same time
physiologically similar GCs of different animals could possess similar morphology
(Yang, Masland, 1994; Kittila, Massey, 1997; MacNeil et al., 1999; Weng et al., 2005).
There are many methods of visualization of retinal neurons: intracellular dye
injection (Yang, Masland, 1994), immunochemistry (Kittila, Massey, 1997), photofilling.
Retrograde tracing with the lipophilic dye from the optic nerve is just convenient to visualize
GCs (Köbbert et al., 2000).
In this study GCs in the fish retina were traced
by the carbocyanine dye DiI applied to the optic nerve cut in retinas preliminary fixed with 4%
paraforme. After 1-2 weeks of incubation in thermostat at 36°C retinal
preparations were investigated under UV microscope. This lipofilic dye diffuses in the lipids of cellular membrane.
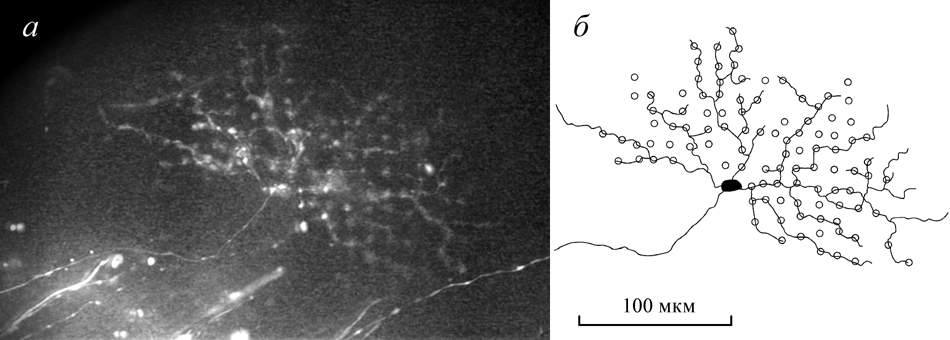
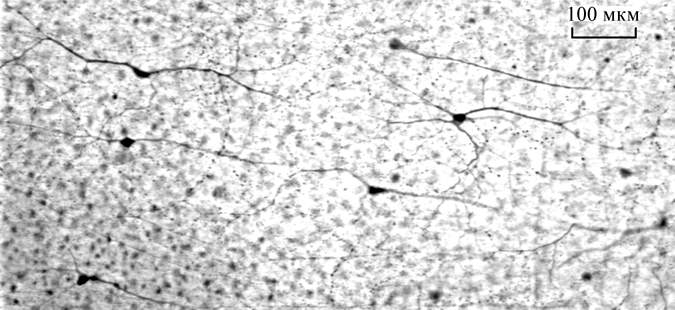
Many axons, cell bodies and completely stained ganglion cell of the pickarel retina traced with DiI are demonstrated in the Fig. 1.
Six types of GCs (among
many other stained) were identified by their clear dendritic morphology.
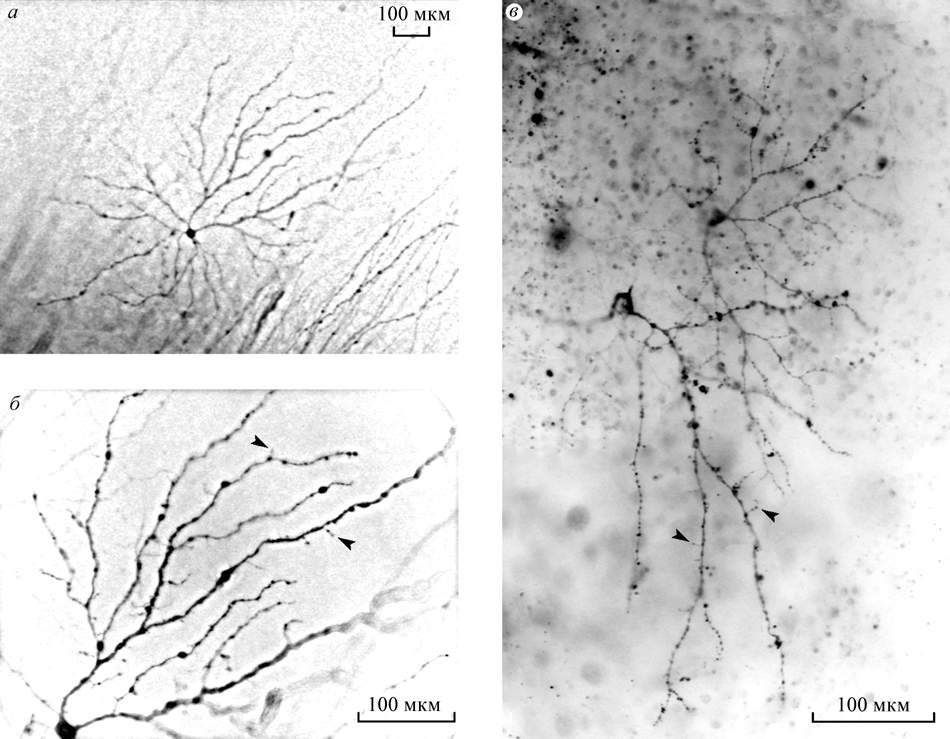
Type 1 and type 2 are GCs of aA and
aB types according to the traditional
morphological classification with large (1500 µ in dia) sparse flat
dendritic arbours stratified in sublaminae a and b of the IPL
correspondingly (Fig.2). In mammalian retinas such cells are classified as
Y cells of off and on physiological types. Type 3 are GCs with dendritic
arborisation situated practically in the same plane as the cell body
similar to d cells of the cat retina (Fig.3 Â).
Those represent so called "slow" direction-selective (DS) GCs projecting to the accessory
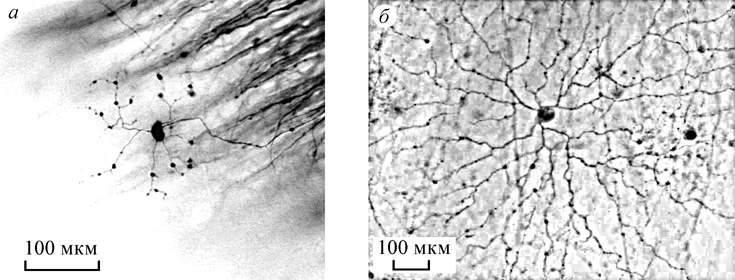
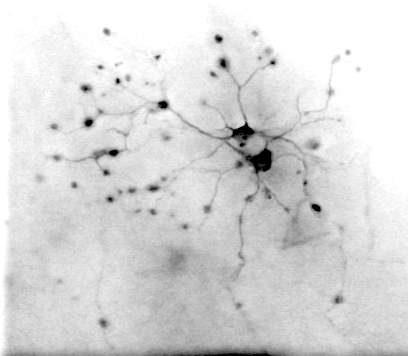
optic system. Type 4 are GCs with small dendritic fields of about 100 µ, their
dendrites are very thin with varicosities at the endings and in
bifurcations (Fig.3 Á). No similar GCs were described in the mammalian
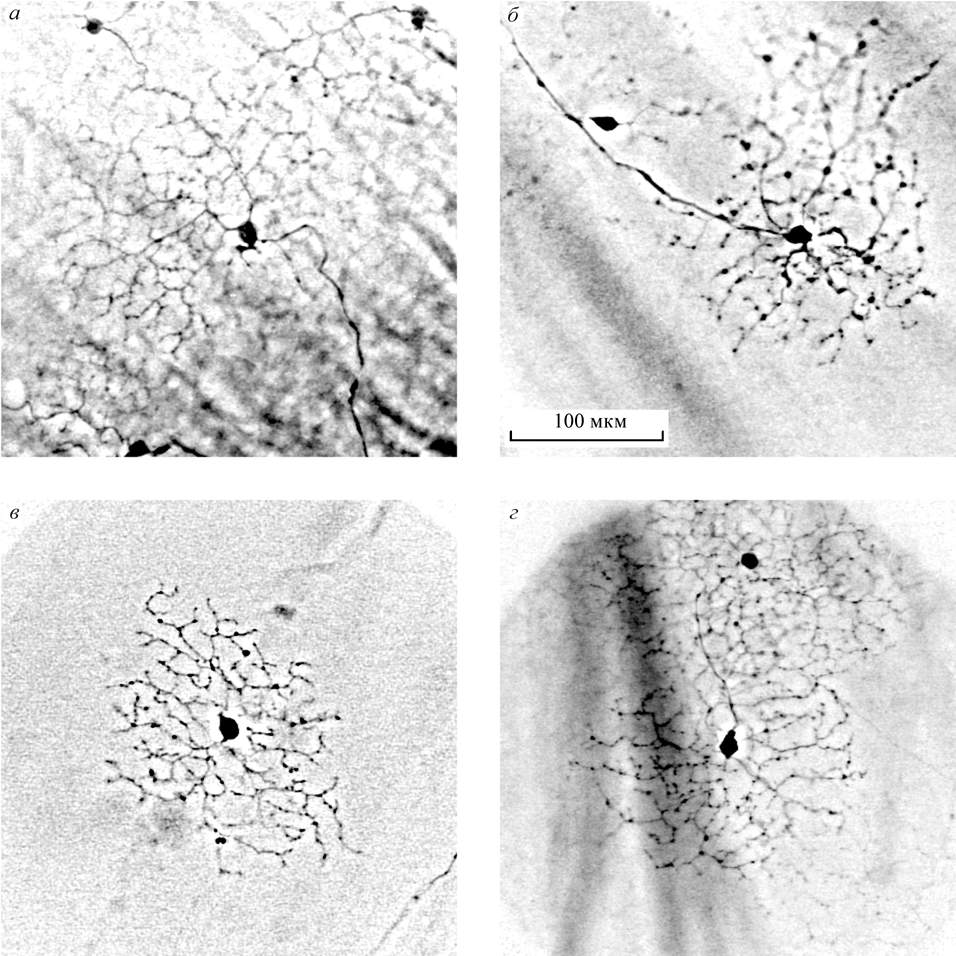
retinas. Type 5 and type 6, these two types of the traced GCs according to
their peculiar dendritic appearance may be identified as "fast" DS GCs (Fig.4).
Their rounded flat "lacy" dendritic arbours are of about 200-300 µ in
diameter. Each type of the putative DS GCs in the fish retina is
monostratified in one of two sublaminae of the IPL. In mammals such "lacy"
dendritic appearance was visualized in the "fast" DS GCs by intracellular dye injection
(Yang, Masland, 1994; Weng et al., 2005). Mammalian DS GCs are bistratified corresponding to
their on-off physiological properties (Kittila, Massey, 1997). On the contrary, there are two
separate (on and off) physiological types of the DS GCs in the fish retina
(Maximov et al., 2005). Thus the revealed essentially monostratified morphology of "lacy"
dendrites is in agreement with the physiological properties of the DS GCs.
Morphological similarities of DS GCs in retinas of different animals imply
common cellular mechanisms of the directional selectivity.
Two types of monostratified starburst amacrine cells are considered to play a critical role in direction selectivity.
However for the fish retina, Djupsund et al. (2003) suggested another candidate for the role of organizer of direction selectivity, namely, a specific type of big fusiform bistratified amacrine cells revealed by different methods
exclusively in the fish retina - Fig. 5.
Maximova EM, Orlov OYu, Dimentman AM (1971) Investigation of visual system of some marine fishes, Voprocy ichtiologii 11: 893-899 (in Russian)
Kittila CA, Massey SC (1997) Pharmacology of directionally selective ganglion cells in the rabbit retina, J Neurophysiol 77: 675-689
Köbbert CC, Apps R, Bechmann I, Lanciego JL, Mey J, Thanos S (2000) Current concepts in neuroanatomical tracing, Progress in Neurobiology 62: 327-351
MacNeil MA, Heussy JK, Dacheux R, Raviola E, Masland RH (1999) The shapes and numbers of amacrine cells: matching of photofilled with Golgi-stained cells in the rabbit retina and comparison with other mammalian species, J Comp Neurol 413: 305-326
Mangrum WI, Dowling JE, Cohen ED (2002) A morphological classification of ganglion cells in the zebrafish retina, Visual Neurosci 19: 767-779
Rockhill RL, Daly FJ, MacNeil MA, Brown SP, Masland RH (2002) The diversity of ganglion cells in a mammalian retina, J Neurosci 22: 3831-3843
Weng S, Sun W, He S (2005) Identification of ON-OFF direction-selective ganglion cells in the mouse retina, J Physiol 562: 915-923
Yang G, Masland RH (1994) Receptive fields and dendritic structure of directionally selective retinal ganglion cells, J Neurosci 14: 5267-5280
Djupsund K, Furukawa T, Yasui S, Yamada M (2003) Asymmetric temporal properties in the receptive field of retinal transient amacrine cells, J Gen Physiol 122: 445-458
Questions and comments to
Back to IITP Electronic Posters Page
Back to IITP Projects Page ( IITP russian Projects Page )
Last Update: June 13, 2006